Функции и строение цитоплазматической мембраны. Наружная цитоплазматическая мембрана, ее строение и функции Какие функции выполняет наружная цитоплазматическая мембрана кратко
Цитоплазматическая мембрана или плазмалемма (лат. membrana – кожица, плёнка) – тончайшая пленка (7– 10нм), отграничивающая внутреннее содержимое клетки от окружающей среды, видна только в электронный микроскоп.
По химической организации плазмалемма представляет липопротеидный комплекс – молекулы липидов и белков .
Её основу составляет липидный бислой, состоящий из фосфолипидов, кроме этого, в мембранах присутствуют гликолипиды и холестерол. Все они обладают свойством амфипатричности, т.е. у них есть гидрофильные («любящие воду») и гидрофобные («боящиеся воды») концы. Гидрофильные полярные «головки» липидных молекул (фосфатная группа) обращены кнаружи мембраны, а гидрофобные неполярные «хвосты» (остатки жирных кислот) – друг к другу, что создает биполярный липидный слой. Молекулы липидов подвижны и могут перемещаться в своем монослое или редко – из одного монослоя в другой. Монослои липидов обладают ассиметричностью, т. е. отличаются по составу липидов, что придает специфичность мембранам даже в пределах одной клетки. Бислой липидов может находиться в состоянии жидкого или твердого кристалла.
Вторым обязательным компонентом плазмалеммы являются белки. Многие мембранные белки способны перемещаться в плоскости мембраны или вращаться вокруг своей оси, но не могут переходить с одной стороны бислоя липидов на другой.
Липиды обеспечивают основные структурные особенности мембраны, а белки – её функции.
Функции мембранных белков различны: поддержание структуры мембран, получение и преобразование сигналов из окружающей среды, транспорт некоторых веществ, катализ реакций, происходящих на мембранах.
Различают несколько моделей строения цитоплазматической мембраны.
①. БУТЕРБРОДНАЯ МОДЕЛЬ (белки – липиды – белки)
В 1935г. английские ученые Даниэли и Даусон высказали идею о послойном расположении в мембранемолекул белков (темные слои в электронном микроскопе), которые залегают снаружи, и молекул липидов (светлый слой) – внутри. Длительное время существовало представление о едином трехслойном строении всех биологических мембран.
При детальном изучении мембраны с помощью электронного микроскопа оказалось, что светлый слой на самом деле представлен двумя слоями фосфолипидов – это билипидный слой , причем водорастворимые его участки – гидрофильные головки направлены к белковому слою, а нерастворимые (остатки жирных кислот) – гидрофобные хвосты обращены друг к другу.
 ②. ЖИДКОСТНО-МОЗАИЧНАЯ МОДЕЛЬ
②. ЖИДКОСТНО-МОЗАИЧНАЯ МОДЕЛЬ
В 1972г .Сингер и Николсон описали модель мембраны, которая получила широкое признание. Согласно этой модели молекулы белков не образуют сплошного слоя, а погружены в биполярный липидный слой на разную глубину в виде мозаики. Глобулы белковых молекул, подобно айсбергам, погружены в «океан»
липидов: одни находятся на поверхности билипидного слоя – периферические белки , другие погружаются в него наполовину – полуинтегральные белки , третьи – интегральные белки – пронизывают его насквозь, формируя гидрофильные поры. Периферические белки, находясь на поверхности билипидного слоя, связаны с головками липидных молекул электростатическими взаимодействиями. Но они никогда не образуют сплошного слоя и, по сути дела, не являются белками собственно мембраны, а, скорее, связывают ее с надмембранной или субмембранной системой поверхностного аппарата клетки.
Основную роль в организации собственно мембраны играют интегральные и полуинтегральные (трансмембранные) белки, имеющие глобулярную структуру и связанные с липидной фазой гидрофильно-гидрофобными взаимодействиями. Молекулы белков, как и липиды, обладают амфипатричностью и своими гидрофобными участками взаимодействуют с гидрофобными хвостами билипидного слоя, а гидрофильные участки обращены к водной среде и образуют с водой водородные связи.
③. БЕЛКОВО-КРИСТАЛЛИЧЕСКАЯ МОДЕЛЬ (модель липопротеинового коврика )
Мембраны образованы переплетением липидных и белковых молекул, объединяющихся между собой на основе гидрофильно-
гидрофобных взаимодействий.
 |
|||

Белковые молекулы, как штифты, пронизывают слой липидов и выполняют в составе мембраны функцию каркаса. После обработки мембраны жирорастворимыми веществами белковый каркас сохраняется, что доказывает взаимосвязь между молекулами белков в мембране. По-видимому, эта модель реализуется лишь в отдельных специальных участках некоторых мембран, где требуется жесткая структура и тесные стабильные взаимоотношения между липидами и белками (например, в области расположения фермента Na-К –АТФ-азы ).
Самой универсальной моделью, отвечающей термодинамическим принципам (принципам гидрофильно-гидрофобных взаимодействий), морфо-биохимическим и экспериментально-цитологическим данным, является жидкостно-мозаичная модель. Однако все три модели мембран не исключают друг друга и могут встречаться в разных участках одной и той же мембраны в зависимости от функциональных особенностей данного участка.
СВОЙСТВА МЕМБРАНЫ
1. Способность к самосборке. После разрушающих воздействий мембрана способна восстановить свою структуру, т.к. молекулы липидов на основе своих физико-химических свойств собираются в биполярный слой, в который затем встраиваются молекулы белков.
2. Текучесть. Мембрана не является жесткой структурой, большая часть входящих в её состав белков и липидов может перемещаться в плоскости мембраны, они постоянно флюктуируют за счет вращательных и колебательных движений. Это определяет большую скорость протекания химических реакций на мембране.
3. Полупроницаемость . Мембраны живых клеток пропускают, помимо воды, лишь определённые молекулы и ионы растворённых веществ. Это обеспечивает поддержание ионного и молекулярного состава клетки.
4. Мембрана не имеет свободных концов . Она всегда замыкается в пузырьки.
5. Асимметричность . Состав наружного и внутреннего слоев как белков, так и липидов различен.
6. Полярность . Внешняя сторона мембраны несёт положительный заряд, а внутренняя – отрицательный.
ФУНКЦИИ МЕМБРАНЫ
1) Барьерная – плазмалемма отграничивает цитоплазму и ядро от внешней среды. Кроме того, мембрана делит внутреннее содержимое клетки на отсеки (компартменты), в которых зачастую протекают противоположные биохимические реакции.
2) Рецепторная (сигнальная) – благодаря важному свойству белковых молекул – денатурации, мембрана способна улавливать различные изменения в окружающей среде. Так, при воздействии на мембрану клетки различных средовых факторов (физических, химических, биологических) белки, входящие в ее состав, меняют свою пространственную конфигурацию, что служит своеобразным сигналом для клетки.
Это обеспечивает связь с внешней средой, распознавание клеток и их ориентацию при формировании тканей и т.д. С этой функцией связана деятельность различных регуляторных систем и формирование иммунного ответа.
3) Обменная – в состав мембраны входят не только структурные белки, которые образуют ее, но и ферментативные, являющиеся биологическими катализаторами. Они располагаются на мембране в виде «каталитического конвейера» и определяют интенсивность и направленность реакций метаболизма.
4) Транспортная – молекулы веществ, диаметр которых не превышает 50 нм, могут проникать путем пассивного и активного транспорта через поры в структуре мембраны. Крупные вещества попадают в клетку путем эндоцитоза (транспорт в мембранной упаковке), требующего затраты энергии. Его разновидностями являются фаго- и пиноцитоз .
Пассивный транспорт – вид транспорта, в котором перенос веществ осуществляется по градиенту химической или электрохимической концентрации без затраты энергии АТФ. Выделяют два вида пассивного транспорта: простая и облегченная диффузия. Диффузия – это перенос ионов или молекул из зоны более высокой их концентрации в зону более низкой концентрации, т.е. по градиенту.
Простая диффузия – ионы солей и вода проникают через трансмембранные белки или жирорастворимые вещества по градиенту концентрации.
Облегченная диффузия – специфические белки-переносчики связывают вещество и переносят его через мембрану по принципу «пинг-понга». Таким способом через мембрану проходят сахара и аминокислоты. Скорость такого транспорта значительно выше, чем простой диффузии. Кроме белков- переносчиков, в облегченной диффузии принимают участие некоторые антибиотики – например, грамитидин и ваномицин.
Поскольку они обеспечивают транспорт ионов, их называют ионофорами .
Активный транспорт – это вид транспорта, при котором расходуется энергия АТФ, он идёт против градиента концентрации. В нем принимают участие ферменты АТФ-азы. В наружной клеточной мембране находятся АТФ-азы, которые осуществляют перенос ионов против градиента концентрации, это явление называется ионным насосом. Примером является натрий-калиевый насос. В норме в клетке больше ионов калия, во внешней среде – ионов натрия. Поэтому по законам простой диффузии калий стремится из клетки, а натрий – в клетку. В противовес этому натрий-калиевый насос накачивает против градиента концентрации в клетку ионы калия, а ионы натрия выносит во внешнюю среду. Это позволяет поддерживать постоянство ионного состава в клетке и её жизнеспособность. В животной клетке одна треть АТФ расходуется на работу натрий-калиевого насоса.
Разновидностью активного транспорта является транспорт в мембранной упаковке – эндоцитоз . Крупные молекулы биополимеров не могут проникать через мембрану, они поступают в клетку в мембранной упаковке. Различают фагоцитоз и пиноцитоз. Фагоцитоз – захват клеткой твердых частиц, пиноцитоз – жидких частиц. В этих процессах выделяют стадии:
1) узнавание рецепторами мембраны вещества; 2) впячивание (инвагинация) мембраны с образованием везикулы (пузырька); 3) отрыв пузырька от мембраны, слияние его с первичной лизосомой и восстановление целостности мембраны; 4) выделение непереваренного материала из клетки (экзоцитоз).
Эндоцитоз является способом питания для простейших. У млекопитающих и человека имеется ретикуло-гистио-эндотелиальная система клеток, способная к эндоцитозу – это лейкоциты, макрофаги, клетки Купфера в печени.
ОСМОТИЧЕСКИЕ СВОЙСТВА КЛЕТКИ
Осмос – односторонний процесс проникновения воды через полупроницаемую мембрану из области с меньшей концентрацией раствора в область с более высокой концентрацией. Осмос обусловливает осмотическое давление.
Диализ – односторонняя диффузия растворенных веществ.
Раствор, в котором осмотическое давление такое же, как и в клетках, называют изотоническим. При погружении клетки в изотонический раствор её объем не изменяется. Изотонический раствор называют физиологическим – это 0,9% раствор хлорида натрия, который широко применяется в медицине при сильном обезвоживании и потери плазмы крови.
Раствор, осмотическое давление которого выше, чем в клетках, называют гипертоническим .
Клетки в гипертоническом растворе теряют воду и сморщиваются. Гипертонические растворы широко применяются в медицине. Марлевая повязка, смоченная в гипертоническом растворе, хорошо впитывает гной.
Раствор, где концентрация солей ниже, чем в клетке, называют гипотоническим . При погружении клетки в такой раствор вода устремляется в нее. Клетка набухает, ее тургор увеличивается, и она может разрушиться. Гемолиз – разрушение клеток крови в гипотоническом растворе.
Осмотическое давление в организме человека в целом регулируется системой органов выделения.
Предыдущая123456789Следующая
ПОСМОТРЕТЬ ЕЩЕ:
Клеточная мембрана также называется плазматической (или цитоплазматической) мембраной и плазмалеммой. Данная структура не только отделяет внутреннее содержимое клетки от внешней среды, но также входит с состав большинства клеточных органелл и ядра, в свою очередь отделяя их от гиалоплазмы (цитозоля) - вязко-жидкой части цитоплазмы. Договоримся называть цитоплазматической мембраной ту, которая отделяет содержимое клетки от внешней среды. Остальными терминами обозначать все мембраны.
Строение клеточной мембраны
В основе строения клеточной (биологической) мембраны лежит двойной слой липидов (жиров). Формирование такого слоя связано с особенностями их молекул. Липиды не растворяются в воде, а по-своему в ней конденсируются. Одна часть отдельно взятой молекулы липида представляет собой полярную головку (она притягивается водой, т. е. гидрофильна), а другая - пару длинных неполярных хвостов (эта часть молекулы отталкивается от воды, т. е. гидрофобна). Такое строение молекул заставляет их «прятать» хвосты от воды и поворачивать к воде свои полярные головки.
В результате образуется двойной липидный слой, в котором неполярные хвосты находятся внутри (обращены друг к другу), а полярные головки обращены наружу (к внешней среде и цитоплазме). Поверхность такой мембраны гидрофильна, а внутри она гидрофобна.

В клеточных мембранах среди липидов преобладают фосфолипиды (относятся к сложным липидам). Их головки содержат остаток фосфорной кислоты. Кроме фосфолипидов есть гликолипиды (липиды + углеводы) и холестерол (относится к стеролам). Последний придает мембране жесткость, размещаясь в ее толще между хвостами остальных липидов (холестерол полностью гидрофобный).
За счет электростатического взаимодействия, к заряженным головкам липидов присоединяются некоторые молекулы белков, которые становятся поверхностными мембранными белками. Другие белки взаимодействуют с неполярными хвостами, частично погружаются в двойной слой или пронизывают его насквозь.
Таким образом, клеточная мембрана состоит из двойного слоя липидов, поверхностных (периферических), погруженных (полуинтегральных) и пронизывающих (интегральных) белков . Кроме того, некоторые белки и липиды с внешней стороны мембраны связаны с углеводными цепями.

Это жидкостно-мозаичная модель строения мембраны была выдвинута в 70-х годах XX века. До этого предполагалась бутербродная модель строения, согласно которой липидный бислой находится внутри, а с внутренней и наружной стороны мембрана покрыта сплошными слоями поверхностных белков. Однако накопление экспериментальных данных опровергло эту гипотезу.
Толщина мембран у разных клеток составляет около 8 нм. Мембраны (даже разные стороны одной) отличаются между собой по процентному соотношению различных видов липидов, белков, ферментативной активности и др. Какие-то мембраны более жидкие и более проницаемые, другие более плотные.
Разрывы клеточной мембраны легко сливаются из-за физико-химических особенностей липидного бислоя. В плоскости мембраны липиды и белки (если только они не закреплены цитоскелетом) перемещаются.
Функции клеточной мембраны
Большинство погруженных в клеточную мембрану белков выполняют ферментативную функцию (являются ферментами). Часто (особенно в мембранах органоидов клетки) ферменты располагаются в определенной последовательности так, что продукты реакции, катализируемые одним ферментом, переходят ко второму, затем третьему и т. д. Образуется конвейер, который стабилизируют поверхностные белки, т. к. не дают ферментам плавать вдоль липидного бислоя.
Клеточная мембрана выполняет отграничивающую (барьерную) от окружающей среды и в то же время транспортную функции. Можно сказать, это ее самое главное назначение. Цитоплазматическая мембрана, обладая прочностью и избирательной проницаемостью, поддерживает постоянство внутреннего состава клетки (ее гомеостаз и целостность).
При этом транспорт веществ происходит различными способами. Транспорт по градиенту концентрации предполагает передвижение веществ из области с их большей концентрацией в область с меньшей (диффузия). Так, например, диффундируют газы (CO 2 , O 2).
Бывает также транспорт против градиента концентрации, но с затратой энергии.
Транспорт бывает пассивным и облегченным (когда ему помогает какой-нибудь переносчик). Пассивная диффузия через клеточную мембрану возможна для жирорастворимых веществ.
Есть особые белки, делающие мембраны проницаемыми для сахаров и других водорастворимых веществ. Такие переносчики соединяются с транспортируемыми молекулами и протаскивают их через мембрану.
3. Функции и строение цитоплазматической мембраны
Так переносится глюкоза внутрь эритроцитов.
Пронизывающие белки, объединяясь, могут образовывать пору для перемещения некоторых веществ через мембрану. Такие переносчики не перемещаются, а образуют в мембране канал и работают аналогично ферментам, связывая определенное вещество. Перенос осуществляется благодаря изменению конформации белка, благодаря чему в мембране образуются каналы. Пример - натрий-калиевый насос.
Транспортная функция клеточной мембраны эукариот также реализуется за счет эндоцитоза (и экзоцитоза). Благодаря этим механизмам в клетку (и из нее) попадают крупные молекулы биополимеров, даже целые клетки. Эндо- и экзоцитоз характерны не для всех клеток эукариот (у прокариот его вообще нет). Так эндоцитоз наблюдается у простейших и низших беспозвоночны; у млекопитающих лейкоциты и макрофаги поглощают вредные вещества и бактерии, т. е. эндоцитоз выполняет защитную функцию для организма.
Эндоцитоз делится на фагоцитоз (цитоплазма обволакивает крупные частицы) и пиноцитоз (захват капелек жидкости с растворенными в ней веществами). Механизм этих процессов приблизительно одинаков. Поглощаемые вещества на поверхности клеток окружаются мембраной. Образуется пузырек (фагоцитарный или пиноцитарный), который затем перемещается внутрь клетки.
Экзоцитоз - это выведение цитоплазматической мембраной веществ из клетки (гормонов, полисахаридов, белков, жиров и др.). Данные вещества заключаются в мембранные пузырьки, которые подходят к клеточной мембране. Обе мембраны сливаются и содержимое оказывается за пределами клетки.
Цитоплазматическая мембрана выполняет рецепторную функцию. Для этого на ее внешней стороне располагаются структуры, способные распознавать химический или физический раздражитель. Часть пронизывающих плазмалемму белков с наружней стороны соединены с полисахаридными цепочками (образуя гликопротеиды). Это своеобразные молекулярные рецепторы, улавливающие гормоны. Когда конкретный гормон связывается со своим рецептором, то изменяет его структуру. Это в свою очередь запускает механизм клеточного ответа. При этом могут открываться каналы, и в клетку могут начать поступать определенные вещества или выводиться из нее.
Рецепторная функция клеточных мембран хорошо изучена на основе действия гормона инсулина. При связывании инсулина с его рецептором-гликопротеидом происходит активация каталитической внутриклеточной части этого белка (фермента аденилатциклазы). Фермент синтезирует из АТФ циклическую АМФ. Уже она активирует или подавляет различные ферменты клеточного метаболизма.
Рецепторная функция цитоплазматической мембраны также включает распознавание соседних однотипных клеток. Такие клетки прикрепляются друг к другу различными межклеточными контактами.
В тканях с помощью межклеточных контактов клетки могут обмениваться между собой информацией с помощью специально синтезируемых низкомолекулярных веществ. Одним из примеров подобного взаимодействия является контактное торможение, когда клетки прекращают рост, получив информацию, что свободное пространство занято.
Межклеточные контакты бывают простыми (мембраны разных клеток прилегают друг к другу), замковыми (впячивания мембраны одной клетки в другую), десмосомы (когда мембраны соединены пучками поперечных волокон, проникающих в цитоплазму). Кроме того, есть вариант межклеточных контактов за счет медиаторов (посредников) - синапсы. В них сигнал передается не только химическим, но и электрическим способом. Синапсами передаются сигналы между нервными клетками, а также от нервных к мышечным.
Клеточная теория
В 1665 г. Р. Гук, рассматривая под микроскопом срез пробки дерева, обнаружил пустые ячейки, которые он назвал "клетками". Он видел только оболочки растительных клеток, и длительное время оболочка считалась основным структурным компонентом клетки. В 1825 г. Я. Пуркине описал протоплазму клеток, а в 1831 г. Р. Броун — ядро. В 1837 г. М. Шлейден пришёл к заключению, что растительные организмы состоят из клеток, и каждая клетка содержит ядро.
1.1. Используя накопившиеся к этому времени данные, Т.
Цитоплазматическая мембрана, ее функции и строение
Шванн в 1839 г. сформулировал основные положения клеточной теории:
1) клетка является основной структурной единицей растений и животных;
2) процесс образования клеток обусловливает рост, развитие и дифференцировку организмов.
В 1858 г. Р. Вирхов — основоположник патологической анатомии — дополнил клеточную теорию важным положением, что клетка может происходить только от клетки (Omnis cellula e cellula) в результате её деления. Он установил, что в основе всех заболеваний лежат изменения структуры и функции клеток.
1.2. Современная клеточная теория включает следующие положения:
1) клетка — основная структурно-функциональная и генетическая единица живых организмов, наименьшая единица живого;
2) клетки всех одноклеточных и многоклеточных организмов сходны по строению, химическому составу и важнейшим проявлениям процессов жизнедеятельности;
3) каждая новая клетка образуется в результате деления исходной (материнской) клетки;
4) клетки многоклеточных организмов специализированы: они выполняют разные функции и образуют ткани;
5) клетка является открытой системой через которую проходят и преобразуются потоки вещества, энергии и информации
Строение и функции цитоплазматической мембраны
Клетка представляет собой открытую саморегулирующуюся систему, через которую постоянно идёт поток вещества, энергии и информации. Эти потоки принимает специальный аппарат клетки, в который входят:
1) надмембранный компонент – гликокаликс;
2) элементарная биологическая мембрана или их комплекс;
3) подмембранный опорно-сократительный комплекс гиалоплазмы;
4) анаболическая и катаболическая системы.
Основной компонент этого аппарата — элементарная мембрана.
Клетка содержит различные типы мембран, но принцип их строения одина-
В 1972 году С. Сингером и Г. Николсоном была предложена жидкостно-мозаичная модель строения элементарной мембраны. Согласно этой модели ее основу также составляет билипидный слой, но белки по отношению к этому слою располагаются по-разному. Часть белковых молекул лежит на поверхности липидных слоев (периферические белки), часть пронизывает один слой липидов (полуинтегральные белки), а часть пронизывает оба слоя липидов (интегральные белки). Липидный слой находится в жидкой фазе ("липидное море"). На наружной поверхности мембран имеется рецепторный аппарат — гликокаликс, образованный разветвлёнными молекулами гликопротеинов, "узнающий" определённые вещества и структуры.
2.3. Свойства мембран: 1) пластичность, 2) полупроницаемость, 3) способность самозамыкаться.
2.4. Функции мембран: 1) структурная — мембрана как структурный компонент входит в состав большинства органоидов (мембранный принцип структуры органоидов); 2) барьерная и регуляторная — поддерживает постоянство химического состава и регулирует все обменные процессы (реакции обмена веществ протекают на мембранах); 3) защитная; 4) рецепторная.
Любая живая клетка отделена от окружающей среды тонкой оболочкой особого строения - цитоплазматической мембраной (ЦПМ). Эукариоты имеют многочисленные внутриклеточные мембраны, отделяющие пространство органелл от цитоплазмы, тогда как для большинства прокариот ЦПМ - это единственная мембрана клетки. У некоторых бактерий и архей она может внедряться внутрь цитоплазмы, образуя выросты и складки различной формы.
ЦПМ любых клеток построены по единому плану и состоят из фосфолипидов (рис. 3.5, а). У бактерий в их состав входят две жирные кислоты обычно с 16-18 атомами углерода в цепочке и с насыщенными или одной ненасыщенной связями, соединенные сложноэфирной связью с двумя гидроксильными группами глицерола. Состав жирных кислот бактерий может варьировать в ответ на изменения окружающей среды, в особенности, температуры. При понижении температуры в составе фосфолипидов увеличивается количество ненасыщенных жирных кислот, что в значительной степени отражается на текучести мембраны. Некоторые жирные кислоты могут быть разветвленными или содержать циклопропановое кольцо. Третья ОН-группа глицерола связана с остатком фосфорной кислоты и через него - с головной группой. Головные группы фосфолипидов могут у разных прокариот иметь разную химическую природу (фосфатидилэта- ноламин, фосфатидилглицерол, кардиолипин, фосфатидилсерин, лецитин и др.), но они устроены проще, чем у эукариот. Например, у Е. coli, они представлены на 75% фосфатидилэтаноламином, на 20% - фосфатидилглицеро- лом, остальные состоят из кардиолипина (дифосфатидилглицерола), фос- фатидилсерина и следовых количеств других соединений. Прочие бактерии имеют более сложные типы мембранных липидов. Некоторые клетки образуют гликолипиды, такие как моногалактозилдиглицерид. Мембранные липиды архей отличаются от эукариотических и бактериальных. Вместо жирных кислот у них присутствуют высшие изопреноидные спирты, прикрепленные к глицеролу простой, а не сложной эфирной связью.
Рис. 3.5.
а - фосфолипида; б - бислойной мембраны
О О о О о о
Такие молекулы составляют мембранный бислой, где гидрофобные части обращены вовнутрь, а гидрофильные - наружу, в окружающую среду и в цитоплазму (рис. 3.5, б). В бислой погружены или пересекают его многочисленные белки, которые могут диффундировать внутри мембраны, иногда образуя сложные комплексы. Мембранные белки имеют ряд важных функций, включая преобразование и запасание метаболической энергии, регуляцию поглощения и выброса всех питательных веществ и продуктов метаболизма. Кроме того, они узнают и передают многие сигналы, отражающие изменения в окружающей среде, и запускают соответствующий каскад реакций, приводящий к клеточному ответу. Такая организация мембран хорошо объясняется жидкокристаллической моделью с мозаичным вкраплением мембранных белков (рис. 3.6).

Рис. 3.6.
Большинство биологических мембран имеют толщину от 4 до 7 нм. Клеточные мембраны хорошо различимы в просвечивающем электронном микроскопе при контрастировании тяжелыми металлами. На электронных микрофотографиях они имеют вид трехслойных образований: два внешних темных слоя показывают положение полярных групп липидов, а светлый средний слой - гидрофобное внутреннее пространство (рис. 3.7).
Другая техника исследования мембран заключается в получении сколов замороженных при температуре жидкого азота клеток и контрастировании образующихся поверхностей с помощью напыления тяжелых металлов
(платина, золото, серебро). Полученные препараты просматривают в сканирующем электронном микроскопе. При этом можно увидеть поверхность мембраны и включенные в нее мозаично мембранные белки, которые не простираются сквозь мембрану, а связаны специальными гидрофобными якорными областями с гидрофобной областью бислоя.

Рис. 3.7.
ЦПМ обладает свойством избирательной проницаемости, препятствуя свободному продвижению большинства веществ внутрь и из клетки, а также играет значительную роль в росте и делении клеток, движении, экспорте поверхностных и внеклеточных белков и углеводов (экзополисахаридов). Если клетку поместить в среду с более высоким или более низким осмотическим давлением, чем внутри цитоплазмы, то произойдет выход воды из клетки или вход воды в нее. Это отражает свойство воды уравнивать градиенты растворов. Цитоплазма при этом сжимается или расширяется (явление плазмолиза/деплазмолиза). Большинство бактерий, однако, не меняет свою форму в таких экспериментах вследствие наличия ригидной клеточной стенки.
ЦПМ регулирует потоки питательных веществ и метаболитов. Наличие гидрофобного слоя, образованного мембранными липидами, препятствует прохождению через нее любых полярных молекул и макромолекул. Это свойство позволяет клеткам, существующим в большинстве случаев в разбавленных растворах, удерживать полезные макромолекулы и метаболические предшественники. Мембрана клетки призвана осуществлять и транспортную функцию. Обычно прокариоты имеют большое количество очень специфических транспортных систем. Транспорт - интегральная часть общей биоэнергетики клетки, которая создает и использует различные ионные градиенты через ЦПМ для переноса веществ и формирования других необходимых клетке градиентов. ЦПМ играет значительную роль в движении, росте и делении клеток. В мембране прокариот сосредоточены многие метаболические процессы. Мембранные белки выполняют важные функции: участвуют в преобразовании и запасании энергии, регулируют поглощение и выброс всех питательных веществ и продуктов метаболизма, узнают и передают сигналы об изменениях в окружающей среде.
Общие сведения об эукариотической клетке
Каждая эукариотическая клетка имеет обособленное ядро, в котором заключен отграниченный от матрикса ядерной мембраной генетический материал (это главное отличие от прокариотических клеток). Генетический материал сосредоточен преимущественно в виде хромосом, имеющих сложное строение и состоящих из нитей ДНК и белковых молекул. Деление клеток происходит посредством митоза (а для половых клеток - мейоза). Среди эукариотов есть как одноклеточные, так и многоклеточные организмы.
Существует несколько теорий происхождения эукариотических клеток, одна из них - эндосимбионтическая. В гетеротрофную анаэробную клетку проникла аэробная клетка типа бактериоподобной, которая послужила базой для появления митохондрий. В эти клетки начали проникать спирохетоподобные клетки, которые дали начало формированию центриолей. Наследственный материал отгородился от цитоплазмы, возникло ядро, появился митоз. В некоторые эукариотические клетки проникли клетки типа сине-зеленых водорослей, которые положили начало появлению хлоропластов. Так впоследствии возникло царство растений.
Размеры клеток тела человека варьируются от 2-7 мкм (у тромбоцитов) до гигантских размеров (до 140 мкм у яйцеклетки).
Форма клеток обусловлена выполняемой ими функцией: нервные клетки - звездчатые за счет большого количества отростков (аксона и дендритов), мышечные клетки - вытянутые, так как должны сокращаться, эритроциты могут менять свою форму при продвижении по мелким капиллярам.
Строение эукариотических клеток животных и растительных организмов во многом схоже. Каждая клетка снаружи ограничена клеточной оболочкой, или плазмалеммой. Она состоит из цитоплазматической мембраны и слоя гликокаликса (толщиной 10-20 нм), который покрывает ее снаружи. Компоненты гликокаликса - комплексы полисахаридов с белками (гликопротеины) и жирами (гликолипиды).
Цитоплазматическая мембрана - это комплекс бислоя фосфолипидов с протеинами и полисахаридами.
В клетке выделяют ядро и цитоплазму . Клеточное ядро стоит из мембраны, ядерного сока, ядрышка и хроматина. Ядерная оболочка состоит из двух мембран, разделенных перинуклеарным пространством, и пронизана порами.
Основу ядерного сока (матрикса) составляют белки: нитчатые, мин фибриллярные (опорная функция), глобулярные, гетероядерные РНК и мРНК (результат процессинга).
Ядрышко - это структура, где происходит образование и созревание рибосомальных РНК (р-РНК).
Хроматин в виде глыбок рассеян в нуклеоплазме и является нитерфазной формой существования хромосом.
В цитоплазме выделяют основное вещество (матрикс, гиалоплазму), органеллы и включения.
Органеллы могут быть общего значения и специальные (в клетках, выполняющих специфические функции: микроворсинки всасывающего эпителия кишечника, миофибриллы мышечных клеток и т. д.).
Органеллы общего значения - эндоплазматическая сеть (гладкая и шероховатая), комплекс Гольджи, митохондрии, рибосомы и полисомы, лизосомы, пероксисомы, микрофибриллы и микротрубочки, центриоли клеточного центра.
В растительных клетках есть еще и хлоропласты, в которых протекает фотосинтез.
Элементарная мембрана состоит из бислоя липидов в комплексе с белками (гликопротеины: белки + углеводы, липопротеины: жиры + белки). Среди липидов можно выделить фосфолипиды, холестерин, гликолипиды (углеводы + жиры), липопротеины. Каждая молекула жира имеет полярную гидрофильную головку и неполярный гидрофобный хвост. При этом молекулы ориентированы так, что головки обращены кнаружи и внутрь клетки, а неполярные хвосты - внутрь самой мембраны. Этим достигается избирательная проницаемость для веществ, поступающих в клетку.
Выделяют периферические белки (они расположены только по внутренней или наружной поверхности мембраны), интегральные (они прочно встроены в мембрану, погружены в нее, способны менять свое положение в зависимости от состояния клетки). Функции мембранных белков: рецепторная, структурная (поддерживают форму клетки), ферментативная, адгезивная, антигенная, транспортная.
Схема строения элементарной мембраны жидкостно-мозаичная: жиры составляют жидкокристаллический каркас, а белки мозаично встроены в него и могут менять свое положение.
Важнейшая функция: способствует компартментации - подразделению содержимого клетки на отдельные ячейки, отличающиеся деталями химического или ферментного состава. Этим достигается высокая упорядоченность внутреннего содержимого любой эукариотической клетки. Компартментация способствует пространственному разделению процессов, протекающих в клетке. Отдельный компартмент (ячейка) представлен какой-либо мембранной органеллой (например, лизосомой) или ее частью (кристами, отграниченными внутренней мембраной митохондрий).
Другие функции:
1) барьерная (отграничение внутреннего содержимого клетки);
2) структурная (придание определенной формы клеткам в соответствии с выполняемыми функциями);
3) защитная (за счет избирательной проницаемости, рецепции и антигенности мембраны);
4) регуляторная (регуляция избирательной проницаемости для различных веществ (пассивный транспорт без затраты энергии по законам диффузии или осмоса и активный транспорт с затратой энергии путем пиноцитоза, эндо- и экзоцитоза, работы натрий-калиевого насоса, фагоцитоза));
5) адгезивная функция (все клетки связаны между собой посредством специфических контактов (плотных и неплотных));
6) рецепторная (за счет работы периферических белков мембраны). Существуют неспецифические рецепторы, которые воспринимают несколько раздражителей (например, холодовые и тепловые терморецепторы), и специфические, которые воспринимают только один раздражитель (рецепторы световоспринимающей системы глаза);
7) электрогенная (изменение электрического потенциала поверхности клетки за счет перераспределения ионов калия и натрия (мембранный потенциал нервных клеток составляет 90 мВ));
8) антигенная: связана с гликопротеинами и полисахаридами мембраны. На поверхности каждой клетки имеются белковые молекулы, которые специфичны только для данного вида клеток. С их помощью иммунная системы способна различать свои и чужие клетки.
Цитоплазматическая мембрана
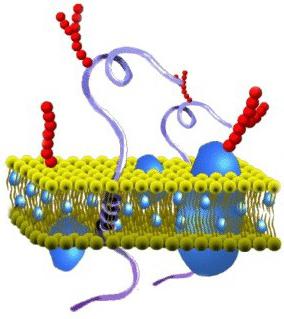
Изображение клеточной мембраны. Маленькие голубые и белые шарики соответствуют гидрофильным «головкам» липидов, а присоединённые к ним линии - гидрофобным «хвостам». На рисунке показаны только интегральные мембранные белки (красные глобулы и желтые спирали). Желтые овальные точки внутри мембраны - молекулы холестерола Желто-зеленые цепочки бусинок на наружной стороне мембраны - цепочки олигосахаридов , формирующие гликокаликс
Биологическая мембрана включает и различные белки : интегральные (пронизывающие мембрану насквозь), полуинтегральные (погруженные одним концом во внешний или внутренний липидный слой), поверхностные (расположенные на внешней или прилегающие к внутренней сторонам мембраны). Некоторые белки являются точками контакта клеточной мембраны с цитоскелетом внутри клетки, и клеточной стенкой (если она есть) снаружи. Некоторые из интегральных белков выполняют функцию ионных каналов, различных транспортеров и рецепторов .
Функции биомембран
- барьерная - обеспечивает регулируемый, избирательный, пассивный и активный обмен веществ с окружающей средой. Например, мембрана пероксисом защищает цитоплазму от опасных для клетки пероксидов . Избирательная проницаемость означает, что проницаемость мембраны для различных атомов или молекул зависит от их размеров, электрического заряда и химических свойств. Избирательная проницаемость обеспечивает отделение клетки и клеточных компартментов от окружающей среды и снабжение их необходимыми веществами.
- транспортная - через мембрану происходит транспорт веществ в клетку и из клетки. Транспорт через мембраны обеспечивает: доставку питательных веществ, удаление конечных продуктов обмена, секрецию различных веществ, создание ионных градиентов, поддержание в клетке соответствующего pH и ионной концентрации, которые нужны для работы клеточных ферментов.
Частицы, по какой-либо причине не способные пересечь фосфолипидный бислой (например, из-за гидрофильных свойств, так как мембрана внутри гидрофобна и не пропускает гидрофильные вещества, или из-за крупных размеров), но необходимые для клетки, могут проникнуть сквозь мембрану через специальные белки-переносчики (транспортеры) и белки-каналы или путем эндоцитоза .
При пассивном транспорте вещества пересекают липидный бислой без затрат энергии, путем диффузии. Вариантом этого механизма является облегчённая диффузия , при которой веществу помогает пройти через мембрану какая-либо специфическая молекула. У этой молекулы может быть канал, пропускающий вещества только одного типа.
Активный транспорт требует затрат энергии, так как происходит против градиента концентрации. На мембране существуют специальные белки-насосы, в том числе АТФаза , которая активно вкачивают в клетку ионы калия (K+) и выкачивают из неё ионы натрия (Na+).
- матричная - обеспечивает определенное взаиморасположение и ориентацию мембранных белков, их оптимальное взаимодействие;
- механическая - обеспечивает автономность клетки, ее внутриклеточных структур, также соединение с другими клетками (в тканях). Большую роль в обеспечение механической функции имеют клеточные стенки, а у животных - межклеточное вещество.
- энергетическая - при фотосинтезе в хлоропластах и клеточном дыхании в митохондриях в их мембранах действуют системы переноса энергии, в которых также участвуют белки;
- рецепторная - некоторые белки, сидящие в мембране, являются рецепторами (молекулами, при помощи которых клетке воспринимает те или иные сигналы).
Например, гормоны, циркулирующие в крови, действуют только на такие клетки-мишени, у которых есть соответствующие этим гормонам рецепторы. Нейромедиаторы (химические вещества, обеспечивающие проведение нервных импульсов) тоже связываются с особыми рецепторными белками клеток-мишеней.
- ферментативная - мембранные белки нередко являются ферментами. Например, плазматические мембраны эпителиальных клеток кишечника содержат пищеварительные ферменты.
- осуществление генерации и проведения биопотенциалов.
С помощью мембраны в клетке поддерживается постоянная концентрация ионов: концентрация иона К+ внутри клетки значительно выше, чем снаружи, а концентрация Na+ значительно ниже, что очень важно, так как это обеспечивает поддержание разности потенциалов на мембране и генерацию нервного импульса .
- маркировка клетки - на мембране есть антигены, действующие как маркеры - «ярлыки», позволяющие опознать клетку. Это гликопротеины (то есть белки с присоединенными к ним разветвленными олигосахаридными боковыми цепями), играющие роль «антенн». Из-за бесчисленного множества конфигурации боковых цепей возможно сделать для каждого типа клеток свой особый маркер. С помощью маркеров клетки могут распознавать другие клетки и действовать согласованно с ними, например, при формировании органов и тканей. Это же позволяет иммунной системе распознавать чужеродные антигены.
Структура и состав биомембран
Мембраны состоят из липидов трёх классов: фосфолипиды , гликолипиды и холестерол . Фосфолипиды и гликолипиды (липиды с присоединёнными к ним углеводами) состоят из двух длинных гидрофобных углеводородных «хвостов», которые связаны с заряженной гидрофильной «головой». Холестерол придаёт мембране жёсткость, занимая свободное пространство между гидрофобными хвостами липидов и не позволяя им изгибаться. Поэтому мембраны с малым содержанием холестерола более гибкие, а с большим - более жёсткие и хрупкие. Также холестерол служит «стопором», препятствующим перемещению полярных молекул из клетки и в клетку. Важную часть мембраны составляют белки, пронизывающие её и отвечающие за разнообразные свойства мембран. Их состав и ориентация в разных мембранах различаются.
Клеточные мембраны часто асимметричны, то есть слои отличаются по составу липидов, переход отдельной молекулы из одного слоя в другой (так называемый флип-флоп ) затруднён.
Мембранные органеллы
Это замкнутые одиночные или связанные друг с другом участки цитоплазмы , отделённые от гиалоплазмы мембранами . К одномембранным органеллам относятся эндоплазматическая сеть , аппарат Гольджи , лизосомы , вакуоли , пероксисомы ; к двумембранным - ядро , митохондрии , пластиды . Снаружи клетка ограничена так называемой плазматической мембраной. Строение мембран различных органелл отличается по составу липидов и мембранных белков.
Избирательная проницаемость
Клеточные мембраны обладают избирательной проницаемостью: через них медленно диффундируют глюкоза , аминокислоты , жирные кислоты , глицерол и ионы , причем сами мембраны в известной мере активно регулируют этот процесс-одни вещества пропускают, а другие нет. существует четыре основных механизма для поступления веществ в клетку или их из клеки наружу:диффузия , осмос , активный транспорт и экзо- или эндоцитоз . Два первых процесса носят пассивный характер, т.е. не требуют затрат энергии; два последних-активные процессы, связанные с потреблением энерги.
Избирательная проницаемость мембраны при пассивном транспорте обусловлена специальными каналами - интегральными белками. Они пронизывают мембрану насквозь, образовывая своего рода проход . Для элементов K, Na и Cl есть свои каналы. Относительно градиента концентрации молекулы этих элементов движутся в клетку и из неё. При раздражении каналы натриевых ионов раскрываются, и происходит резкое поступление в клетку ионов натрия. При этом происходит дисбаланс мембранного потенциала. После чего мембранный потенциал восстанавливается. Каналы калия всегда открыты, через них в клетку медленно попадают ионы
Наружная цитоплазматическая мембрана представляет собой тончайшую пленку. Ее толщина - порядка 7-10 нм. Просматривается пленка только в электронный микроскоп.
Структура
Какой состав имеет цитоплазматическая мембрана? Строение пленки достаточно разнообразно. В соответствии с химической организацией, она представляет собой комплекс белков и липидов. Цитоплазматическая мембрана клетки включает в себя бислой. Он выступает в качестве основы. Кроме этого, цитоплазматическая мембрана содержит холестерол и гликолипиды. Этим веществам свойственна амфипатричность. Другими словами, в них присутствуют гидрофобные ("боящиеся влаги") и гидрофильные ("любящие воду") концы. Последние (фосфатная группа) направлены наружу от мембраны, вторые (остатки от жирных кислот) ориентированы друг к другу. За счет этого и формируется липидный биполярный слой. Липидные молекулы обладают подвижностью. Они способны перемещаться в собственном монослое либо (что редко) из одного в другой.
Липидный слой может иметь состояние твердого или жидкого кристалла. Монослои отличаются асимметричностью. Это значит, что в них различен состав липидов. За счет этого свойства цитоплазматические мембраны обладают специфичностью даже в рамках одной клетки. Ко второму обязательному компоненту пленки относят белки. Многие из этих соединений могут перемещаться в мембранной плоскости либо совершать вращения вокруг собственной оси. При этом они не способны переходить из одной части бислоя в другую. Защита внутренней среды - основная задача, которую выполняет цитоплазматическая мембрана. Строение пленки, кроме этого, обеспечивает течение различных процессов. За выполнение тех или иных задач отвечают белки. Благодаря липидам обеспечиваются структурные особенности пленки.
Цитоплазматическая мембрана: функции
Основными задачами являются:
- Барьерная . Защитная пленка обеспечивает активный, пассивный, избирательный, регулируемый обмен соединений с внешней средой. За счет избирательной проницаемости осуществляется отделение клетки и ее компартментов и снабжение их нужными веществами.
- Транспортная . Сквозь пленку осуществляется переход соединений от клетки к клетке. Благодаря этому доставляются питательные соединения, удаляются конечные продукты обмена, происходит секреция разных веществ. Кроме этого, формируются ионные градиенты, на оптимальном уровне поддерживаются ионная концентрация и рН. Они необходимы для активной деятельности ферментов клетки.
Вспомогательные задачи

Особые свойства
К специфическим функциям мембраны относят:

Дополнительные сведения
Если какие-то частицы по тем или другим причинам не способны пройти сквозь фосфолипидный бислой (к примеру, вследствие гидрофильных свойств, поскольку внутри цитоплазматическая мембрана гидрофобна и такие соединения не пропускает, либо из-за больших размеров самих частиц), но они необходимы, то пройти они могут с помощью специальных белков-переносчиков (транспортеров) и белков-каналов. Либо проникновение их осуществляется посредством эндоцитоза.

В процессе пассивного транспорта пересечение веществами липидного слоя происходит путем диффузии. При этом энергия не затрачивается. В качестве одного из вариантов такого механизма может выступать облегченная диффузия. В ходе нее облегчает прохождение вещества какая-нибудь специфическая молекула. У нее может присутствовать канал, способный пропускать только однотипные частицы. При активном транспорте затрачивается энергия. Это связано с тем, что данный процесс осуществляется против концентрационного градиента. Цитоплазматическая мембрана содержит особые белки-насосы, АТФазу в том числе, которая способствует активному вхождению калиевых и выведению натриевых ионов.
Модели
Их существует несколько:
- "Бутербродная модель" . Идею о трехслойном строении всех мембран высказали ученые Даусон и Даниэли в 1935 году. По их мнению, структура пленки была следующей: белки-липиды-белки. Такое представление существовало достаточно долго.
- "Жидкостно-мозаичная структура". Эта модель была описана Николсоном и Сингером в 1972 году. В соответствии с ней белковые молекулы не формируют сплошной слой, а погружаются в биполярный липидный в виде мозаики на различную глубину. Эта модель считается наиболее универсальной.
- "Белково-кристаллическая структура". В соответствии с этой моделью мембраны формируются за счет переплетения белковых и липидных молекул, которые объединены на базе гидрофильно-гидрофобных связей.